




Abstract
MicroRNAs (miRNAs) are a class of small non-coding RNAs that have emerged as potential predictive, prognostic, and therapeutic biomarkers, relevant to many pathophysiological conditions including limb immobilization, osteoarthritis, sarcopenia, and cachexia. Impaired musculoskeletal homeostasis leads to distinct muscle atrophies. Understanding miRNA involvement in the molecular mechanisms underpinning conditions such as muscle wasting may be critical to developing new strategies to improve patient management. MicroRNAs are powerful post-transcriptional regulators of gene expression in muscle and, importantly, are also detectable in the circulation. MicroRNAs are established modulators of muscle satellite stem cell activation, proliferation, and differentiation, however, there have been limited human studies that investigate miRNAs in muscle wasting. This narrative review summarizes the current knowledge as to the role of miRNAs in the skeletal muscle differentiation and atrophy, synthesizing the findings of published data.
Cite this article: Bone Joint Res 2020;9(11):798–807.
Article focus
-
Accumulating evidence suggests a significant role of microRNA (miRNA) in muscle differentiation and an important role of miRNAs as regulators of muscle atrophy.
-
This review summarizes the role of miRNAs in skeletal muscle differentiation and atrophy and offers future research directions.
-
MicroRNAs are involved in muscle atrophies associated with different pathophysiological conditions.
Key messages
-
MicroRNAs regulate myogenesis, through the control of satellite cell quiescence, proliferation, and terminal differentiation.
-
There is no consensus on how miRNA dysregulation alters the pattern of muscle wasting related to different pathophysiologies.
-
More research is required to understand the involvement of miRNAs in muscle atrophy pathways before these targets can be used as clinical biomarkers.
Strengths and limitations
-
Potential specific miRNAs are suggested as possible markers and therapeutic targets for muscle-related diseases, which can be considered for future studies.
-
Although the actual details of how specific miRNAs affect muscle-wasting mechanisms related to different pathophysiologies are limited, the current knowledge concerning which miRNAs are involved in each of the major disease area is summarized.
-
In the future, as the literature in this field expands, more targeted, systematic reviews would be useful to explore detailed facets of the muscle miRNA literature, such as specific markers of the onset of muscle wasting following immobilization.
Introduction
Skeletal muscle is critical for health throughout life. Loss of muscle mass is typically seen with ageing and occurs in many chronic diseases including cancer cachexia, osteoarthritis (OA), and type II diabetes mellitus. It can also present itself more acutely through disuse atrophy, evident as a result of bed rest and limb immobilization. All muscle atrophy conditions share common functional consequences, through the loss of muscle strength and power, reducing the ability to perform activities of daily living, with a resultant impact on a person’s quality of life. These are major causes of morbidity and are poor prognostic factors for many patients.1 The pathogenesis of muscle wasting appears to be multifactorial, involving inflammation, impaired muscle regeneration, oxidative stress, senescence, and apoptosis.2,3 Studies suggest that these complex cellular processes play a role in myofibre degeneration followed by satellite cell activation and differentiation.2,3 The muscle regenerative response consists of activation of quiescent satellite cells, proliferation of the myogenic precursor cells (myoblasts), and terminal differentiation into myocytes and their fusion into myofibres. However, failed myofibre regeneration and/or accelerated pathological processes that affect cellular homeostasis eventually lead to muscle atrophy.2,3 Therefore, an understanding of the formation of muscle (myogenesis), but also an understanding of molecular mechanisms underpinning muscle wasting, is critical to the development of prognostic or therapeutic biomarkers.
MicroRNAs (miRNAs) belong to a class of small non-coding RNA species that post-transcriptionally alter gene expression by increasing translational repression or by inducing messenger RNA (mRNA) cleavage and degradation.4,5 They are involved in a variety of biological processes through their regulatory function in mechanisms of post-transcriptional gene expression (Table I).6-10 Cell and animal studies have demonstrated that miRNAs are altered during muscle cell proliferation, differentiation, apoptosis, and regeneration.2,3,6 In recent years, miRNAs have emerged as regulators of skeletal muscle function and aberrant expression of miRNAs is thought to be associated with the progression of muscle atrophy in various diseases.11 For example, miR-29b has been reported to be commonly upregulated in multiple types of muscle atrophy.12 miRNAs have been shown to play an important role in human embryonic myogenesis13 and also in adult myogenesis after injury.14 Individual miRNAs have been shown to be involved in myogenesis, through the control of satellite cell quiescence (miR-195, miR-497), proliferation (miR-133, miR-27), and additionally myoblast differentiation (miR-206, miR-1, miR-486).15-19 In addition, miRNAs are secreted into extracellular fluids. Circulating miRNAs are attractive potential biomarkers for a variety of muscle atrophy conditions, which can also serve as signalling molecules to mediate intracellular communications. As such, these may be attractive prognostic and therapeutic biomarkers, which could serve as adjunct tools in the management of patients with muscle atrophies. This review describes the biogenesis and function of miRNAs, and highlights their role in skeletal muscle differentiation and regulation of muscle atrophy. The format of our review offers a perspective on the role of miRNAs in diseases related to muscle dysfunction investigated in different models. Due to a relatively low number of available publications, our study has used a narrative review format, which has allowed us to provide a “best-evidence synthesis” and relevant interpretation of the data. More specifically, the limitations, which determined a narrative review format of this paper, result from: 1) small number of publications for specific miRNAs for particular muscle conditions in cohort studies; 2) discrepancies between in vitro, animal, and in vivo data; and 3) additional difficulty related to categorization of investigated models and confounding factors between them. As such, although this review is not exhaustive, it can serve as a guide for future work in this area.
Table I.
MicroRNAs expressed in muscular tissue and their global effect on muscle biological process and their validated targets.
| miR | Biological process | Tissue expression | Validated mRNA targets | Reference |
|---|---|---|---|---|
| miR-1 | enhancer of skeletal muscle differentiation | muscle-specific | HDAC4, Pax7, Pax3, |
15–17,20,21 |
| miR-133a/b | enhancer of myoblast proliferation | muscle-specific | FGFR1, PP2AC, PRDM16, SRF, IGF1 | 15 |
| miR-206 | enhancer of myoblast proliferation | muscle-specific | Notch3, IGFß, Meox2, RARB, Fzd7, MAPK3, CLCN3, NFAT5, Mstn, Smad3, Pax7 | 16,17,20,21 |
| miR-27 a/b | promotes entry into differentiation programme | ubiquitous | Mstn, Pax3 | 16,17,22 |
| miR-26a | promotes myoblast differentiation | ubiquitous | Smad1, Smad4, Ccnd1, Ezh2 | 23 |
| miR-221/222 | promote cell cycle progression | ubiquitous | p27 | 22 |
| miR-146b | promotes satellite cell differentiation | ubiquitous | Smad4, Notch1, Hmga2 | 24 |
| miR-26a | promotes myoblast differentiation | ubiquitous | TGFß/BMP, Smad4 | 23 |
| miR-155 | represses myoblast differentiation | ubiquitous | TNF-α, Cdc25A | 25 |
| miR-503, miR-322/424 | promote myogenesis interfering with the progression through the cell cycle | ubiquitous | TNF-α, Cdc25A | 25 |
| miR-29b | enhancer of skeletal muscle atrophy | ubiquitous | Murf-1, Atrogin-1 | 12 |
-
BMP, bone morphogenetic protein 2; Cdc, cell division cycle; CLCL3, chloride voltage-gated channel 3; HDAC4, histone deacetylase 4; Hmga2, high mobility group at-hook 2; IGF, insulin-like growth factor; MAPK, mitogen-activated protein k ; miRNA, microRNA; mRNA, messenger RNA; Mstn, myostatin; Notch1, notch receptor 1; RARB, retinoic acid receptor beta; SRF, serum response factor; TGFß, tumour growth factor-beta; TNF-α, tumour necrosis factor-alpha.
The sources used for articles in this narrative review were PubMed and MEDLINE. They were searched using the keywords: ‘miRNA in skeletal muscle’, ‘miRNA role in atrophy and regeneration’, ‘miRNA in cancer cachexia’, ‘miRNA in sarcopenia’, ‘miRNA in bed immobilization’, ‘miRNA in OA’, and ‘miRNA in osteoporosis’. The abstracts were reviewed and all that were relevant to the topic of our study were selected. Additionally, the quality of the publications was assessed based on the rank of the journal, and was further aided in some cases by taking into account the number of citations.
MicroRNA biogenesis and function
MicroRNAs are transcribed in the nucleus by the RNA polymerase II enzyme to produce a primary-miRNA transcript (pri-miRNAs), which can be several hundred base pairs in length.26 Pri-miRNAs form specific hairpin secondary structures and enter a microprocessor complex, which contains ribonuclease III enzyme Drosha, which cleaves pri-miRNAs at the 3' and 5' end, leaving an approximately 70-nucleotide stem-loop precursor miRNA (pre-miRNA). Pre-miRNAs are exported from the nucleus to the cytoplasm via EXPORTIN 5, a nuclear transport protein, which recognizes double-stranded RNAs. In the cytoplasm, pre-miRNAs are further processed by the endoribonuclease Dicer to a short double-strand miRNA duplex, leaving a short double-stranded RNA duplex, which harbours the mature miRNA. The double-stranded RNA duplex unwinds and then the mature single-stranded miRNA associates with the RNA-induced silencing complex (RISC). This RISC consists of multiple proteins27 and when it binds with the miRNA it forms the miRNA-induced silencing complex (miRISC). As part of the miRISC, miRNAs can then interact with the untranslated region 3′ (3′UTR) of target messenger RNAs (mRNAs) and either inhibit their translation or induce mRNA cleavage and degradation (Figure 1).
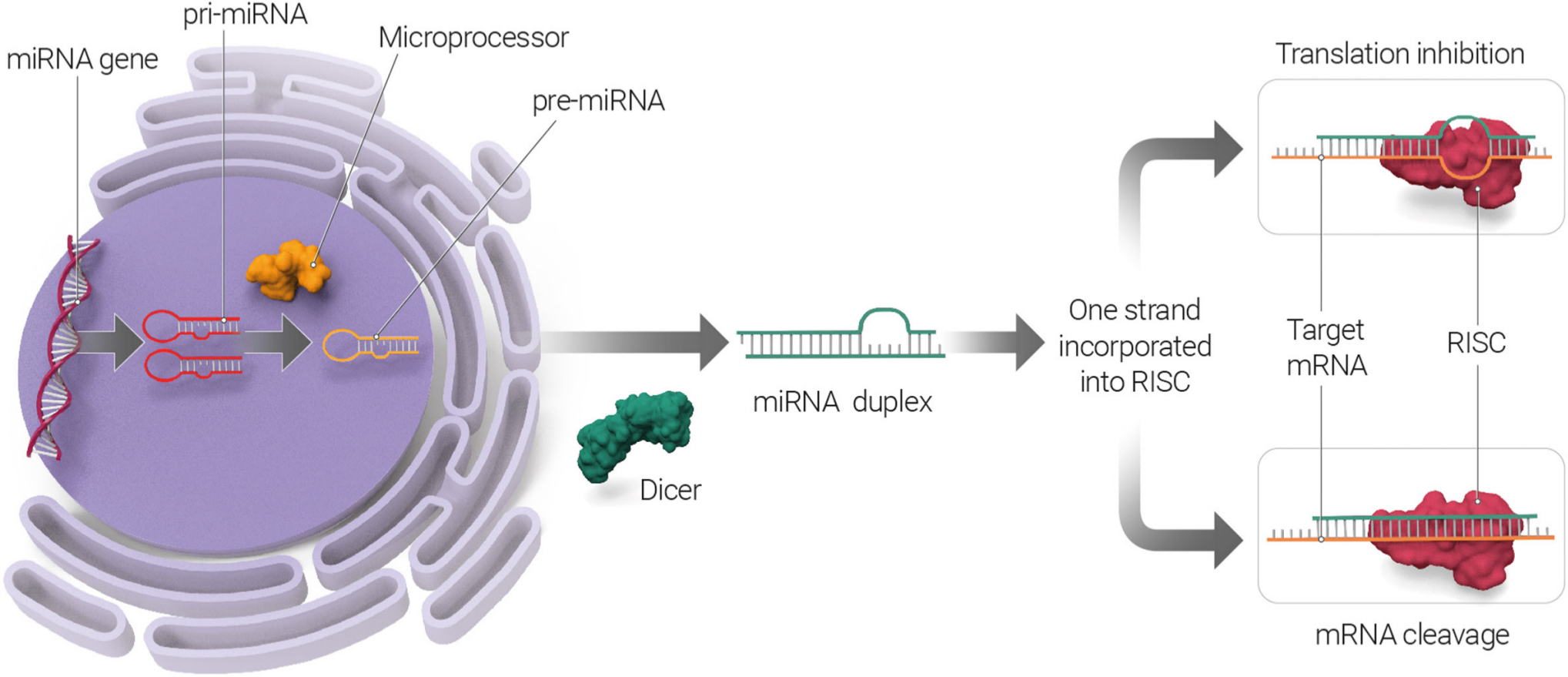
Fig. 1
MicroRNA (miRNA) biogenesis pathway. miRNAs are transcribed in the nucleus, generating primary miRNAs (pri-miRNAs) that undergo nuclear cleavage to form precursor miRNAs (pre-miRNAs). In the cytoplasm, pre-miRNAs are further processed by the Dicer. The double-stranded RNA duplex unwinds and then the mature single-stranded miRNAs assemble into RNA-induced silencing complex (RISC). The miRNA targets messenger RNA (mRNA) to either inhibit mRNA translation or induce mRNA cleavage and degradation.
Muscle stem cells
Skeletal muscle has a high regenerative capacity in response to injury and exercise. This regenerative potential is fulfilled by myogenic precursors known as satellite stem cells (SSCs), which express characteristic paired-box transcription factors, such as Pax7 and Pax3.28–30 SSCs reside beneath the basal lamina, closely juxtaposed to the plasma membrane, and are mitotically quiescent in adults.31 They are activated in response to stimulation (such as muscle injury or exercise) by re-entering the cell cycle. This results in satellite cell proliferation and myofibre regeneration. It has been demonstrated that during this time SSCs encounter different fates (Figure 2). The majority are a fast-dividing population of Pax7+MyoD+ cells that undergo limited replication before myogenesis and are committed to differentiation, whereas a minor population of slow-dividing Pax7+MyoD– cells revert to quiescent self-renewing cells.32,33
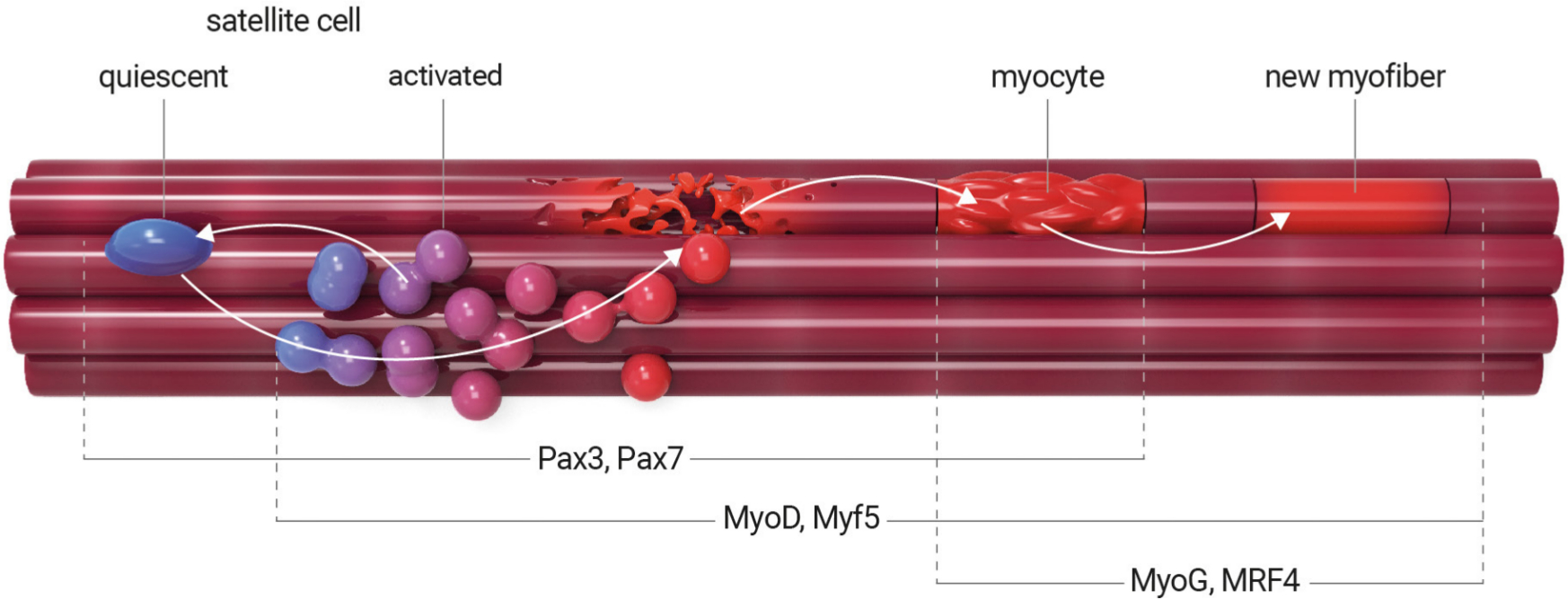
Fig. 2
Satellite stem cell differentiation process. In healthy adult muscle, satellite cells remain in a nonproliferative, quiescent state. They are activated in response to muscle injury or exercise. Activated satellite cells proliferate, undergo self-renewal, and differentiate into myoblasts and then to myocytes, which can mutually fuse and generate myotubes. During this time, the satellite stem cells encounter different fates. Quiescent satellite cells are characterized by the expression of transcription factors Pax7 and Pax3, whereas activated satellite cells coexpress Pax7 and myogenic differentiation factors Myf5 and MyoD. Activation of MyoG and Mrf4 characterizes the terminally differentiated myocyte. Diseases can impair satellite cell activation and proliferation, resulting in the inhibition of terminal differentiation.
Role of miRNA in controlling skeletal muscle differentiation
Myogenesis is an ordered multi-step process during which activated SSCs stop proliferating, undergo differentiation, and fuse into myotubes (Figure 2).20 This involves a complex of transcriptional mediators but also non-coding RNA molecules. Satellite cells maintain the myogenic regulatory factors (MRFs), which are required for myogenic determination. Primary MRFs, such as myogenic differentiation (MyoD) and myogenic factor 5 (Myf5), act as determination genes, which initiate an early stage of myogenesis and characterize proliferating progenitor cells, known as myoblasts. Secondary MRFs, myogenin (MyoG) and myogenic factor 6 (Mrf4), are specific to late-stage myogenesis and characterize the terminally differentiated myocyte. In addition, the MRF family interacts synergistically with myocyte enhancer factor 2 (Mef2) to activate skeletal muscle-specific transcription promoters. Expression and splicing changes of Mef2 proteins isoforms Mef2A, Mef2C, and Mef2D in response to MyoD are consistent with the observation that the majority of skeletal muscle genes require both MyoD and Mef2 family members to activate myogenesis. Therefore, Mef2 appears to play a specific role in early events of cell differentiation. The whole process is followed by fusion into regenerating fibres. Disruption of this network entirely abrogates skeletal muscle formation, regenerative potential, and remodelling.
Recent studies have incorporated miRNAs into the complex myogenicregulatory network. Within skeletal muscle, the 'muscle-specific' miRNAs (e.g. miR-1, miR-133a, and miR-206) play a central role in muscle biology and are all induced during differentiation of myoblasts into myotubes. However, regulation between myogenic transcription factors and miRNAs is complex and depends on the cell cycle and fusion stages. For instance, overexpression of miR-1 or miR-206 in myoblasts leads to accelerated differentiation into myotubes accompanied by a decrease in cell proliferation.17-19 The overexpression of miR-1 leads to decreased levels of histone deacetylase 4 (HDAC4) protein in C2C12 cells,17 and the overexpression of HDAC4 has been demonstrated to repress both myoblast differentiation and Mef2 dependent transcription.21 Conversely, overexpression of miR-133 promotes myoblast proliferation and decreases serum response factor (SRF) levels, thus preventing SRF-mediated inhibition of myoblast proliferation.17 MicroRNAs also play a modulatory role in the upstream regulation of muscle stem cell factors such as Pax3 and Pax7; miR-1, miR-27, and miR-206 all inhibit Pax3,18,19 whereas miR–1 and miR-206 repress Pax722,34 and thus promote the terminal differentiation of myoblasts downstream of MyoD. However, the inhibition of miR-27b maintains Pax3 expression and delays differentiation. While some miRNAs are upregulated during the first stage of differentiation, others, such as miR-221 and miR-222, are downregulated,24 which is correlated with elevated expression of the cell cycle inhibitor p27 (the target of both miR-221 and miR-222). Overexpression of miR-221 and miR-222 delays cell cycle withdrawal and differentiation.24
More recently, an abundance of miRNA-489 has been identified in quiescent satellite cells and it has been shown that this is quickly downregulated during satellite cell activation, while satellite cells that lack a functional miRNA pathway spontaneously exit the quiescent stage and enter the cell cycle.23 Furthermore, miR-195 and miR-497 have been shown to play a role in the maintenance of quiescence in juvenile muscle stem cells, by targeting the cell cycle genes, cell division cycle 25 (Cdc25), and cyclin d2 (Ccnd).15 These findings are particularly relevant to the development of muscle stem cell therapies, as manipulation of these miRNAs prior to transplantation may prompt more efficient regeneration of muscle. A novel miR-146b has also been found among the miRNAs that are upregulated during satellite cell activation and myoblast differentiation. It was proposed that its upregulation is accompanied by downregulation of the target genes such as Smad family member 4 (Smad4), notch receptor 1 (Notch1), and high mobility group at-hook 2 (Hmga2) in order to allow the activation of myogenic differentiation programme.25 It is believed that miR-26a and miR-146b act together to regulate Smad4 during myogenesis. In vitro cell culture and animal studies established that miR-26a promotes muscle differentiation and regeneration by downregulating, signalling pathways that inhibit differentiation, (TGF-ß/BMP) as well as smad family member 1 and 4 (Smad1, Smad4), by targeting specifically 39 3′ UTRs.35 A recent study has not only established the role of miR-26a for skeletal muscle development in vivo but also its regulatory role in muscle differentiation and regeneration through myogenesis repression.35
Furthermore, recent in vitro and in vivo (rodent) studies have shown that miR-27a/b plays a role in: 1) the activation of satellite cells and their self-renewal; 2) myoblast proliferation; and 3) accelerated muscle regeneration through negative regulation of myostatin (Mstn).16,17 The loss of Mstn activates satellite cells, increasing muscle growth and regeneration, whereas its increased level leads to muscle atrophy through protein degradation and blocked protein synthesis.16 Associated with the loss of Mstn, there is myoblast activation with an increased number of Pax7 and MyoD positive cells, followed by miR-27a activation due to loss of Mstn. On the other hand, it has also been shown that miR-27 downregulates PAX3 protein levels without affecting the level of PAX7.18 However, Mstn has also been shown to be a negative regulator of Pax7 expression, therefore the increased number of Pax7 positive cells are more likely due to miR-27-mediated inhibition of Mstn rather than to direct regulation of Pax7 by miR-27. Furthermore, in vivo skeletal muscle treatment with miR-27a/b-specific antago-miRs has been shown to result in significant reduction in the numbers of Pax7+cells and activated myoblasts (MyoD+), which further confirms that miR-27 is capable of Mstn regulation.16 Satellite stem cell impairment is consistent with Smad3 loss associated with increased levels of Mstn and decreased levels of miR-27, reported in Smad3-null mice. It has been shown that treatment with Mstn increased miR-27a/b expression, which was dependent on the activity of Smad3. Taken together, this evidence indicates that there is an autoregulatory mechanism in which Mstn, via Smad3 signalling, regulates miR-27a/b expression, which is responsible for satellite activation during muscle regeneration.16 To date several miRNAs have been identified that play a role in muscle stem cell activation and myogenesis, which offer the future prospect of being able to activate myogenesis directly. However, it will be important for further work to provide evidence that manipulation of these miRNAs can reduce or reverse muscle injury and muscle wasting in pre-clinical models.
Role of miRNA in skeletal muscle in 'in vitro' models
Inflammatory cytokine-induced muscle atrophy
The atrophic loss of muscle mass can be triggered by tumour necrosis factor (TNF-α) and insulin-like growth factor (IGF) that regulate myogenesis through complex signalling pathways that affect muscle gene expression. The physiological role of TNF-α links nuclear factor kappa B (NF-κB) with its inhibitory effect on muscle differentiation through the negative regulation of myogenic regulatory factors (MRFs). Conversely, IGF activates NF-κB, which increases both myoblast proliferation and differentiation. A recent in vitro study showed that the TNF-α inhibitory effect on muscle myogenesis is mediated by regulatory miRNAs.36 A small number of miRNAs are regulated by TNF-α or IGF1. TNF-α stimulates expression of miR-155, which is repressed during myoblast differentiation, but overexpression of miR-155 inhibits myotube formation. TNF-α induces miR-503 downregulation during myoblast differentiation whereas miR-503 expression is upregulated during myotube formation. Parallel inhibition of miR-155 and overexpression of miR-503 protects against the inhibitory effect of TNF-α on myotube formation. It is postulated that intervention at the miRNA level may reduce the inhibitory effect of proinflammatory TNF-α and thereby indirectly induce myoblast differentiation. TNF-α induced suppression of myoblast differentiation was demonstrated to be relieved by overexpression of miR-1, miR-206, and miR-133a/b in the murine C2C12 cell-line.37 This beneficial effect can be considered as an innovative therapeutic strategy in the context of skeletal muscle atrophy. It has been suggested that biogenesis of some miRNAs is modulated by both TNF-α and IGF1 via mitogen-activated protein k (MAPK) signalling pathway and that this might have therapeutic significance.36 TNF-like weak inducer of apoptosis (TWEAK) is an inflammatory cytokine, which belongs to the TNF super ligand family, but is more potent than its structural counterpart TNF-α. TWEAK treatment of C2C12 myotubes resulted in the inhibition of miR-1 to 1, miR-1 to 2, miR-133a, miR-133b, and miR-206, while miR-146a and miR-455 were increased.38 Collectively the gene targets of miRNAs modulated by TWEAK are involved in the inflammatory response, fibrosis, extracellular matrix remodelling, and proteolytic degradation.38
Dexamethasone-induced muscle atrophy
Dexamethasone is a corticosteriod used for the treatment of a various inflammatory-related conditions, as well as being an adjunctive treatment in cancer patients undergoing chemotherapy. Dexamethasone is widely used in in vitro muscle systems to induce muscle atrophy. A number of studies have reported that miRNAs play a role in dexamethasone-induced atrophy in cultured muscle cells.39-41 High-throughput analysis of miRNAs using microarrays indicates there are multiple candidate miRNAs which are altered by dexamethasone-induced atrophy.39 Dexamethasone treatment of C2C12 myotubes led to the upregulation of 11 miRNAs and downregulation of six miRNAs. Independent validation by stem-loop real-time quantitative reverse transcription polymerase chain reaction (RT-PCR) confirmed six miRNAs were differentially expressed including miR-1, miR-147, miR-332, miR-351, miR-503, and miR-708. The targets of these miRNAs include many muscle growth and atrophy-related genes.39 Another study showed that transfection of miR-182 into C2C12 myotubes treated with dexamethasone significantly suppressed forkhead box O3 (FoxO3) transcriptional factors at the mRNA and protein level (Figure 3). Furthermore, transfection of miR-182 which targets the 3′ UTR of FoxO3, also resulted in the downregulation of multiple targets which are transcriptionally regulated by FoxO3, including F-box protein coding genes (Mafbx and Atrogin-1) among others.40 Intriguingly, extracellular miR-182 levels were increased in the cell culture media following dexamethasone treatment of C2C12 myotubes,41 which suggests miR-182 may be actively or passively released during glucocorticoid-induced atrophy.
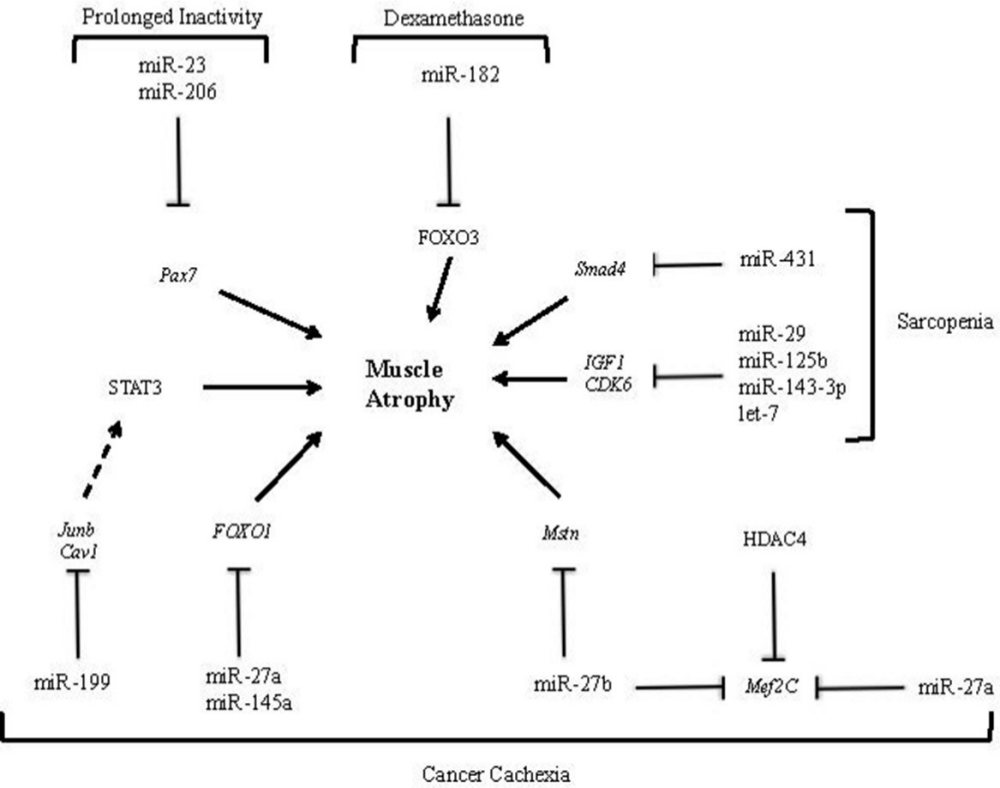
Fig. 3
The involvement of microRNAs (miRNAs) in muscle atrophy associated with cancer cachexia, sarcopenia, prolonged inactivity, and dexamethasone-induced atrophy. MicroRNA-23 and miR-206 downregulation affects the expression of transcription factor Pax7 in satellite stem cells, promoting muscle mass wasting during prolonged activity.42 Following dexamethasone treatment, miR-182 suppresses forkhead box O3 (FOXO3) at the messenger RNA (mRNA) and protein level, which induces atrophy.40 In sarcopenia, miR-29, miR-125b, miR-143-3p, and let-7 have been shown to impair myogenesis by targeting insulin-like growth factor 1 (IGF-1) and cyclin-dependent kinase 6 (CDK6).43 In mice muscle, miR-431 was overexpressed and the muscle has shown reduced Smad4 level, which increases during ageing.44 Interactions between miRNA and mRNA have been identified in cancer cachexia muscle wasting. They include miR-199a/caveolin 1 (Cav1), miR-199a/transcription factor Jun-B (Junb), miR-27a/FOXO1, miR-145a/FOXO1, miR-27a/myocyte enhancer factor 2 (Mef2C), and miR-27b/Mef2C.45 STAT3, signal transducer and activator of transcription 3.
Role of miRNA in immobilization
Localized immobilization-induced muscle atrophy
Regular mechanical loading is essential for the maintenance of muscle mass and function. Limb immobilization or hind limb suspension leads to localized skeletal muscle atrophy. This model represents changes within muscles, which may appear after the conservative treatment of long bone fractures using plaster casts. High-throughput profiling of miRNAs in the soleus muscle after two to seven days hind limb suspension in an animal model revealed changes in 18 miRNAs. Reduced miR-499 and miR-208b were identified as potential regulators of the muscle atrophy. Both of these markers were reduced even further after longer periods (observed up to 28 days) of hind limb suspension. Conversely, expression of miR-499, which targets SRY-box transcription factor 6 (Sox6), was increased. Sox6 target is established as negative regulators of ß-myosin heavy chain (ß-MHC), which encodes the contractile protein of the myosin heavy chain.46 Another study identified that the miR-30 family was downregulated by hind limb suspension. Similar changes in the miR-30 family were observed in dystrophic Mdx4cv compared to wild-type mice. SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily D member 2 (Smarcd2), snail family transcriptional repressor 2 (Snai2), and trinucleotide repeat containing adaptor 6a (Tnrc6a) were validated as miR-30 family targets, which may indirectly modulate other myogenic miRNAs such as miR-206.47 Muscle unloading also occurs under zero gravity conditions: one previous study revealed that 11 days of spaceflight resulted in transcriptional modulation of several hundred genes, notably miR-206 expression in gastrocnemius was found to be significantly decreased.48
Bed rest-induced muscle atrophy
Bed rest and lack of all physical activity in comparison with limb immobilization can induce well-recognized systemic metabolic changes such as decreased cardiovascular function, decreased muscle function, and increased insulin resistance.49 This model can be used to mimic physiological changes and the consequences of sustained inactivity and unloading, which occurs in some patients after trauma. Analysis of the vastus lateralis muscle in six males after ten days of bed rest showed downregulation of miR-206, a factor responsible for muscle function maintenance, and also a downregulation of miR-23a, a factor responsible for insulin messaging and defence against muscle atrophy (Figure 3).42 However, it has been shown that resistance training induces miR-206 upregulation and thus activates SSC proliferation, enhancing muscle differentiation.50 Downregulation of miR-23 shows an overlap with the dexamethasone-treated patients where the expression of this miRNA was also decreased, highlighting the role of this miRNA in muscle homeostasis.48 Rezen et al42 has shown that several members of the let-7 family, which affects the cell cycle and insulin resistance, were also downregulated. However, another study on vastus lateralis muscle samples, after a 21-day bed rest period, showed that the let-7 family of miRNAs was upregulated. This miRNA group is increased in type II diabetes mellitus and is associated with inactivity.51 Furthermore, the let-7 group is upregulated in muscle samples from the elderly, suggesting an overlap of the mechanisms involved in the response to unloading and ageing.52 Patients in intensive care units (ICUs) are characterized by long-term immobilization, bed rest, and artificial ventilation, leading to accelerated muscle wasting. Respiratory muscle is particularly susceptible to wasting, when artificial ventilation is necessary. One clinical study has assessed miRNA expression in muscle of septic ICU patients, which reported an increase in primary miR-21 levels, but not in its mature form of miR-21, which suggests that impairment of miR-21 processing may occur in muscles of ICU patients.53 This group of patients represents both bed rest-induced as well as cytokine-induced muscle atrophy.
Role of miRNA in skeletal muscle atrophy
Cancer cachexia
Cancer cachexia is a complex syndrome that affects patients with advanced cancer and impacts adversely on quality of life, morbidity, and mortality.54 In cancer patients, there is a wide variation in muscle atrophy and some patients appear to be more susceptible to cachexia. New interactions between miRNA and mRNA have been identified in cancer cachexia muscle wasting. These include miR-27a/FOXO1, miR-27a/Mef2C, miR-27b/Cxcl12, miR-27b/Mef2C, miR-140/Cxcl12, miR-199a/caveolin 1 (Cav1), and miR-199a/Junb.45 MicroRNA-27a and miR-27b have been shown to regulate Mef2, the gene responsible for activation of skeletal muscle-specific transcription promoters during muscle regeneration.45,55 MicroRNA-27b has also been shown to downregulate myostatin (Mstn), a member of the transforming growth factor β (TGFβ) family, responsible for SSC activation and myoblast proliferation. Jiang et al56 have demonstrated that an increase in miR-145a decreased forkhead box O1 (FOXO1) expression, suggesting that miR-145a should be considered as a potential regulator during muscle decline in cancer cachexia. In addition, miR-199 has been linked to Cav1 and transcription factor Jun-B (Junb) regulation. Junb regulates gene expression on multiple levels, however it has been shown to interact with signal transducer and activator of transcription 3 (STAT3), which contributes to cancer cachexia (Figure 3).57 High-throughput sequencing of small RNAs in skeletal muscle from cachexia patients may further help to reveal novel therapeutic targets within muscle. However, muscle itself may also be a source of extracellular miRNAs. Microvesicles provide intracellular transport for different proteins and miRNAs, which are present in many different tissues. A study on miRNAs transported in microvesicles revealed the presence of miR-21, which was secreted by tumour cells, and which induced concomitant myoblast cell death.58 Microvesicles harbouring miR-21 were proposed to fuse with muscle cells and to activate toll like receptor (TLR7/8), leading to apoptosis.58 In this respect, repeated measurements of circulating miRNAs over several timepoints in cancer patients would be valuable for the discovery of novel miRNA biomarkers which can track muscle wasting.
Sarcopenia
In adult humans, muscle mass is lost at a rate of ~1%/year after the age of 30 years.59 This process is thought to decrease regenerative capacity in the muscle.2 Sarcopenia is defined as an age-related muscle decline. It develops by itself and often progresses independently of coexisting comorbidities that are associated with the chronic inflammatory state.60 The role of miRNA in ageing muscle is complex and confounded by factors such as physical activity. A total of 26 miRNAs have been identified to be regulated by age and exercise, but the interaction of these factors has also been shown to affect miRNA expression.61 Investigation of the role of miRNAs in sarcopenic patients has shown up regulation of both has-miR-34a-5p and has-miR-449b-5p. These miRNAs significantly increased the expression of a key senescence gene sirtuin 1 (SIRT1) and other genes related to the mitogen-activated protein kinase (MAPK) pathway, which regulate ageing processes.62 Additionally, IGF-1 and cell cycle regulators such as cyclin-dependent kinase 6 (CDK6) are downregulated in ageing muscles. MicroRNA-29, miR-125b, miR-143 to 3 p, and let-7 have been shown to impair myogenesis by targeting IGF-1 and CDK6 (Figure 3).43
Similarly, transforming growth factor β (TGF-β) gene has been shown to play an important part in sarcopenia and to be excessively activated in skeletal muscle ageing.44 It has been demonstrated that miR-26a, miR-675, miR-146b, and miR-431 inhibit TGF-β signalling and improve muscle regeneration. Interestingly, when miR-431 was overexpressed in mice, it improved myogenic capacity of myoblasts. Likewise, when it was injected into mice muscle, the muscle has shown reduced Smad family member 4 (Smad4) levels, which increase during ageing (Figure 3). In conclusion, this study highlighted a possible role of miR-431 as a therapeutic target in muscle ageing.44
Role of miRNA in related disorders
Osteoporosis
Osteoporosis is the most common metabolic bone disease, characterized by decreasing bone quantity over time, leading to decreased bone strength. Like sarcopenia, osteoporosis is an age-related disease and there is a growing consensus that the two diseases share common pathways including reduced anabolic hormone sensitivity, increased cytokine activity, and possibly even miRNA expression patterns.63 The role of miRNA in bone homeostasis and their effect on osteoblast or osteoclast differentiation, function, and apoptosis has been established. Some miRNAs (including miR-21 and miR-148) have been suggested as biomarkers for early diagnosis of osteoporosis. However, no specific diagnostic or predictive miRNA has proven to be efficient in clinical practice.64 As discussed, miR-21 has been shown to be upregulated in muscles of immobilized ICU patients and it has been shown to be a miRNA of interest in cancer cachexia.58 These results suggest that miR-21 plays an important role in homeostasis of the musculoskeletal system, however more research is required to understand its specific function in distinct muscle atrophies.
Osteoarthritis
Osteoarthritis (OA) is a multifactorial disease characterized by joint degeneration and pathological changes in musculoskeletal tissues such as the synovium, cartilage, and ligaments. Chondrocytes in OA are dysregulated, producing matrix-degrading enzymes. In short, inflammatory and biomechanical stress signalling pathways are similar and they induce and overexpress cytokines and chemokine genes. It remains controversial whether inflammatory mediators are primary or secondary regulators of cartilage damage in OA, however biomechanical stress, inflammatory changes, and muscle changes may all be linked and influenced by miRNAs.65,66 Periarticular loss of muscle bulk and function is well reported in OA, however the pathway and cross-communication between the OA joints and their effect on muscle is not well understood. MicroRNA-143 is differentially expressed in the cartilage of the elderly suffering with OA.67 Likewise, miR-143 was demonstrated to regulate senescence of satellite cells and adult muscle stem cells via insulin-like growth factor binding protein 5 (IGFBP5), suggesting a link between OA and muscle.68 Similarly, miR-181 was downregulated in sarcopenic muscle samples69 and a further study has demonstrated that the inhibition of this miRNA attenuates OA in animal models.70 Another miRNA, which suggests a link between muscle metabolism and OA, is the downregulation of miR-378 in elderly patients. This specific miRNA is responsible for muscle metabolism, myogenesis, and autophagy.68 Synovial samples taken from late-stage OA joints have identified changes in miR-378 expression. Interestingly, miR-378 has been proven to be a pro-bone regeneration molecule.71 These results cumulatively suggest that miR-378 can be considered as a factor involved in musculoskeletal homeostasis and a link between OA and the associated muscle changes. Importantly, miRNA analysis has shown that the function of miRNA is context-dependent and therefore a question arises whether miRNA-associated mechanisms of muscle wasting are similar or different in OA.72 To date many individual miRNAs have been identified to influence the local pathogenesis within the synovium and cartilage, affecting the expression of signalling molecules including tumour necrosis factor (TNF-α), interleukin 1 (IL-1), and interleukin 6 (IL-6), however apart from the discussed associations, no clear miRNA has been identified as a systemic biomarker predicting muscular phenotype or functional status of patients.73
In conclusion, there is accumulating evidence, from both in vitro and preclinical in vivo studies, that miRNAs play a key role in muscle differentiation and are important regulators of muscle atrophy. This narrative review highlights that there is no consensus on how miRNA dysregulation alters the specific muscle wasting related to different pathophysiologies and may serve as a guide for future studies in this area. There are some promising candidate miRNAs, miR-21 and miR-431, identified in preclinical studies, which now need to be confirmed in human muscle disorder clinical trials. However, interventional clinical trials investigating putative miRNA drugs have exhibited significant efficacy in a range of other health conditions, including hepatitis, pathological fibrosis, cancer, and kidney disease.74 Specifically, the therapeutic effect of miR-29 was studied in keloid and scar tissue formation, and miR-155 was investigated for treatment of T cell lymphoma, clearly suggesting a viable future for miRNA therapeutic agents in various diseases.74 Considering the notable role of miR-2912 and miR-15536 in the muscle, the two miRNA drugs highlight a therapeutic potential of these biomarkers for muscle-related atrophy and dysfunction.
Current research has not outlined a definitive role of specific miRNA in muscle health and disease. The majority of studies evaluating muscle decline are performed in animal (quadrupedal) models. Therefore, the discrepancies of published data do not take into account the differences in studied species, skeletal muscle biopsy sites, age of studied animals, and the physiological as well as locomotor kinematic differences between quadrupedal animals and bipedal species (humans). Moreover, the time course of miRNA changes, with the development and progression of muscle atrophy, has not been determined in a longitudinal study. To date most work compares miRNAs identified in atrophy to healthy muscle, which reflect the consequences of, rather than the cause, of atrophy.
Future studies should also consider the complicated nature of miRNAs and that confounding factors can alter up- or downregulation of these targets. In order to understand better how miRNAs affect expression and function of muscle genes, further work is needed to investigate the mechanisms which control the biogenesis of miRNA. RNA-binding proteins have been identified as the upstream modulators of miRNA, however more research is required to identify these proteins, their role on miRNA, and the extracellular cross talk between different organs that induce their action. The majority of miRNA research has identified specific miRNA targets, which are dysregulated in human disease, however to implement preventative therapies we need to better understand the processes which lead to the miRNA dysregulation in the first place. A clearer understanding of this process may offer the opportunity to identify key targets in disease pathogenesis for predictive and therapeutic biomarkers. Finally, through translational medicine research more effort needs to be placed on developing novel, sensitive, and specific clinical tests that identify the deregulated miRNA biomarkers in a patient’s blood and tissue biopsies.
References
1. Bokshan SL , DePasse JM , Daniels AH . Sarcopenia in orthopedic surgery . Orthopedics . 2016 ; 39 ( 2 ): e295 – e300 . Crossref PubMed Google Scholar
2. Brzeszczyńska J , Johns N , Schilb A , et al. Loss of oxidative defense and potential blockade of satellite cell maturation in the skeletal muscle of patients with cancer but not in the healthy elderly . Aging . 2016 ; 8 : 1690 – 1702 . Google Scholar
3. Brzeszczyńska J , Meyer A , McGregor R , et al. Alterations in the in vitro and in vivo regulation of muscle regeneration in healthy ageing and the influence of sarcopenia . J Cachexia Sarcopenia Muscle . 2018 ; 9 ( 1 ): 93 – 105 . Crossref PubMed Google Scholar
4. Lau NC , Lim LP , Weinstein EG , Bartel DP . An abundant class of tiny RNAs with probable regulatory roles in Caenorhabditis elegans . Science . 2001 ; 294 ( 5543 ): 858 – 862 . Crossref PubMed Google Scholar
5. Lee RC , Ambros V . An extensive class of small RNAs in Caenorhabditis elegans . Science . 2001 ; 294 ( 5543 ): 862 – 864 . Crossref PubMed Google Scholar
6. Takahara S , Lee SY , Iwakura T , et al. Altered expression of microRNA during fracture healing in diabetic rats . Bone Joint Res . 2018 ; 7 ( 2 ): 139 – 147 . Crossref PubMed Google Scholar
7. Wang C , Zheng G-F , Xu X-F . Microrna-186 improves fracture healing through activating the bone morphogenetic protein signalling pathway by inhibiting Smad6 in a mouse model of femoral fracture: an animal study . Bone Joint Res . 2019 ; 8 ( 11 ): 550 – 562 . Crossref PubMed Google Scholar
8. Yu CD , Miao WH , Zhang YY , Zou MJ , Yan XF . Inhibition of miR-126 protects chondrocytes from IL-1β induced inflammation via upregulation of Bcl-2 . Bone Joint Res . 2018 ; 7 ( 6 ): 414 – 421 . Crossref PubMed Google Scholar
9. Zhao R , Wang S , Jia L , Li Q , Qiao J , Peng X . Interleukin-1 receptor antagonist protein (IL-1ra) and miR-140 overexpression via pNNS-conjugated chitosan-mediated gene transfer enhances the repair of full-thickness cartilage defects in a rabbit model . Bone Joint Res . 2019 ; 8 ( 3 ): 165 – 178 . Crossref PubMed Google Scholar
10. Li M , Zhang C , Yang Y . Effects of mechanical forces on osteogenesis and osteoclastogenesis in human periodontal ligament fibroblasts: A systematic review of in vitro studies . Bone Joint Res . 2019 ; 8 ( 1 ): 19 – 31 . Crossref PubMed Google Scholar
11. Güller I , Russell AP , Aaron PR . Micrornas in skeletal muscle: their role and regulation in development, disease and function . J Physiol . 2010 ; 588 ( Pt 21 ): 4075 – 4087 . Crossref PubMed Google Scholar
12. Li J , Chan MC , Yu Y , et al. miR-29B contributes to multiple types of muscle atrophy . Nat Commun . 2017 ; 8 ( 1 ): 1 – 15 . Crossref PubMed Google Scholar
13. O'Rourke JR , Georges SA , Seay HR , et al. Essential role for Dicer during skeletal muscle development . Dev Biol . 2007 ; 311 ( 2 ): 359 – 368 . Crossref PubMed Google Scholar
14. Cheung TH , Quach NL , Charville GW , et al. Maintenance of muscle stem-cell quiescence by microRNA-489 . Nature . 2012 ; 482 ( 7386 ): 524 – 528 . Crossref PubMed Google Scholar
15. Sato T , Yamamoto T , Sehara-Fujisawa A . miR-195/497 induce postnatal quiescence of skeletal muscle stem cells . Nat Commun . 2014 ; 5 : 4597 . Crossref PubMed Google Scholar
16. McFarlane C , Vajjala A , Arigela H , et al. Negative auto-regulation of myostatin expression is mediated by Smad3 and microRNA-27 . PLoS One . 2014 ; 9 ( 1 ): 87687 . Crossref PubMed Google Scholar
17. Chen J-F , Mandel EM , Thomson JM , et al. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation . Nat Genet . 2006 ; 38 ( 2 ): 228 – 233 . Crossref PubMed Google Scholar
18. Crist CG , Montarras D , Pallafacchina G , et al. Muscle stem cell behavior is modified by microRNA-27 regulation of Pax3 expression . Proc Natl Acad Sci U S A . 2009 ; 106 ( 32 ): 13383 – 13387 . Crossref PubMed Google Scholar
19. Hirai H , Verma M , Watanabe S , Tastad C , Asakura Y , Asakura A . Myod regulates apoptosis of myoblasts through microRNA-mediated down-regulation of Pax3 . J Cell Biol . 2010 ; 191 ( 2 ): 347 – 365 . Crossref PubMed Google Scholar
20. Berkes CA , Tapscott SJ . Myod and the transcriptional control of myogenesis . Semin Cell Dev Biol . 2005 ; 16 ( 4-5 ): 585 – 595 . Crossref PubMed Google Scholar
21. Miska EA , Langley E , Wolf D , Karlsson C , Pines J , Kouzarides T . Differential localization of HDAC4 orchestrates muscle differentiation . Nucleic Acids Res . 2001 ; 29 ( 16 ): 3439 – 3447 . Crossref PubMed Google Scholar
22. Chen J-F , Tao Y , Li J , et al. Microrna-1 and microRNA-206 regulate skeletal muscle satellite cell proliferation and differentiation by repressing Pax7 . J Cell Biol . 2010 ; 190 ( 5 ): 867 – 879 . Crossref PubMed Google Scholar
23. Cheung TH , Quach NL , Charville GW , et al. Maintenance of muscle stem-cell quiescence by microRNA-489 . Nature . 2012 ; 482 ( 7386 ): 524 – 528 . Crossref PubMed Google Scholar
24. Cardinali B , Castellani L , Fasanaro P , et al. Microrna-221 and microrna-222 modulate differentiation and maturation of skeletal muscle cells . PLoS One . 2009 ; 4 ( 10 ): e7607 . Crossref PubMed Google Scholar
25. Khanna N , Ge Y , Chen J . Microrna-146B promotes myogenic differentiation and modulates multiple gene targets in muscle cells . PLoS One . 2014 ; 9 ( 6 ): e100657 . Crossref PubMed Google Scholar
26. Bartel DP . Micrornas: target recognition and regulatory functions . Cell . 2009 ; 136 ( 2 ): 215 – 233 . Crossref PubMed Google Scholar
27. Ha M , Kim VN . Regulation of microRNA biogenesis . Nat Rev Mol Cell Biol . 2014 ; 15 ( 8 ): 509 – 524 . Crossref PubMed Google Scholar
28. Beauchamp JR , Heslop L , Yu DS , et al. Expression of CD34 and myf5 defines the majority of quiescent adult skeletal muscle satellite cells . J Cell Biol . 2000 ; 151 ( 6 ): 1221 – 1234 . Crossref PubMed Google Scholar
29. Kuang S , Chargé SB , Seale P , Huh M , Rudnicki MA . Distinct roles for Pax7 and Pax3 in adult regenerative myogenesis . J Cell Biol . 2006 ; 172 ( 1 ): 103 – 113 . Crossref PubMed Google Scholar
30. Relaix F , Montarras D , Zaffran S , et al. Pax3 and Pax7 have distinct and overlapping functions in adult muscle progenitor cells . J Cell Biol . 2006 ; 172 ( 1 ): 91 – 102 . Crossref PubMed Google Scholar
31. Mauro A . Satellite cell of skeletal muscle fibers . J Biophys Biochem Cytol . 1961 ; 9 : 493 – 495 . Crossref PubMed Google Scholar
32. Shinin V , Gayraud-Morel B , Gomès D , Tajbakhsh S . Asymmetric division and cosegregation of template DNA strands in adult muscle satellite cells . Nat Cell Biol . 2006 ; 8 ( 7 ): 677 – 682 . Crossref PubMed Google Scholar
33. Conboy MJ , Karasov AO , Rando TA . High incidence of non-random template strand segregation and asymmetric fate determination in dividing stem cells and their progeny . PLoS Biol . 2007 ; 5 : e102 . Crossref PubMed Google Scholar
34. Dey BK , Gagan J , Dutta A . miR-206 and -486 induce myoblast differentiation by downregulating Pax7 . Mol Cell Biol . 2011 ; 31 ( 1 ): 203 – 214 . Crossref PubMed Google Scholar
35. Dey BK , Gagan J , Yan Z , Dutta A . miR-26A is required for skeletal muscle differentiation and regeneration in mice . Genes Dev . 2012 ; 26 ( 19 ): 2180 – 2191 . Crossref PubMed Google Scholar
36. Meyer SU , Thirion C , Polesskaya A , et al. TNF-α and IGF1 modify the microRNA signature in skeletal muscle cell differentiation . Cell Commun Signal . 2015 ; 13 : 4 . Crossref PubMed Google Scholar
37. Georgantas RW , Streicher K , Greenberg SA , et al. Inhibition of myogenic microRNAs 1, 133, and 206 by inflammatory cytokines links inflammation and muscle degeneration in adult inflammatory myopathies . Arthritis Rheumatol . 2014 ; 66 ( 4 ): 1022 – 1033 . Crossref PubMed Google Scholar
38. Panguluri SK , Bhatnagar S , Kumar A , et al. Genomic profiling of messenger RNAs and microRNAs reveals potential mechanisms of TWEAK-induced skeletal muscle wasting in mice . PLoS One . 2010 ; 5 ( 1 ): e8760 . Crossref PubMed Google Scholar
39. Shen H , Liu T , Fu L , et al. Identification of microRNAs involved in dexamethasone-induced muscle atrophy . Mol Cell Biochem . 2013 ; 381 ( 1-2 ): 105 – 113 . Crossref PubMed Google Scholar
40. Hudson MB , Rahnert JA , Zheng B , Woodworth-Hobbs ME , Franch HA , Price SR . miR-182 attenuates atrophy-related gene expression by targeting FOXO3 in skeletal muscle . Am J Physiol Cell Physiol . 2014 ; 307 ( 4 ): C314 – C319 . Crossref PubMed Google Scholar
41. Hudson MB , Woodworth-Hobbs ME , Zheng B , et al. miR-23A is decreased during muscle atrophy by a mechanism that includes calcineurin signaling and exosome-mediated export . Am J Physiol Cell Physiol . 2014 ; 306 ( 6 ): C551 – C558 . Crossref PubMed Google Scholar
42. Režen T , Kovanda A , Eiken O , Mekjavic IB , Rogelj B . Expression changes in human skeletal muscle miRNAs following 10 days of bed rest in young healthy males . Acta Physiol . 2014 ; 210 ( 3 ): 655 – 666 . Crossref PubMed Google Scholar
43. Jung HJ , Lee K-P , Kwon K-S , Suh Y . Micrornas in skeletal muscle aging: current issues and perspectives . J Gerontol A Biol Sci Med Sci . 2019 ; 74 ( 7 ): 1008 – 1014 . Crossref PubMed Google Scholar
44. Lee K-P , Shin YJ , Panda AC , et al. miR-431 promotes differentiation and regeneration of old skeletal muscle by targeting Smad4 . Genes Dev . 2015 ; 29 ( 15 ): 1605 – 1617 . Crossref PubMed Google Scholar
45. Freire PP , Fernandez GJ , Cury SS , et al. The pathway to cancer cachexia: MicroRNA-Regulated networks in muscle wasting based on integrative meta-analysis . Int J Mol Sci . 2019 ; 20 ( 8 ): 1962 . Crossref PubMed Google Scholar
46. McCarthy JJ , Esser KA , Peterson CA , Dupont-Versteegden EE . Evidence of myomiR network regulation of beta-myosin heavy chain gene expression during skeletal muscle atrophy . Physiol Genomics . 2009 ; 39 ( 3 ): 219 – 226 . Crossref PubMed Google Scholar
47. Guess MG , Barthel KKB , Harrison BC , Leinwand LA . miR-30 family microRNAs regulate myogenic differentiation and provide negative feedback on the microRNA pathway . PLoS One . 2015 ; 10 ( 2 ): e0118229 . Crossref PubMed Google Scholar
48. Allen DL , Bandstra ER , Harrison BC , et al. Effects of spaceflight on murine skeletal muscle gene expression . J Appl Physiol . 2009 ; 106 ( 2 ): 582 – 595 . Crossref PubMed Google Scholar
49. Rudrappa SS , Wilkinson DJ , Greenhaff PL , Smith K , Idris I , Atherton PJ . Human skeletal muscle disuse atrophy: effects on muscle protein synthesis, breakdown, and insulin Resistance-A qualitative review . Front Physiol . 2016 ; 7 : 361 . Crossref PubMed Google Scholar
50. D'Souza RF , Markworth JF , Aasen KMM , Zeng N , Cameron-Smith D , Mitchell CJ . Acute resistance exercise modulates microRNA expression profiles: combined tissue and circulatory targeted analyses . PLoS One . 2017 ; 12 ( 7 ): e0181594 – 15 . Crossref PubMed Google Scholar
51. Jiang LQ , Franck N , Egan B , et al. Autocrine role of interleukin-13 on skeletal muscle glucose metabolism in type 2 diabetic patients involves microRNA let-7 . Am J Physiol Endocrinol Metab . 2013 ; 305 ( 11 ): E1359 – E1366 . Crossref PubMed Google Scholar
52. Drummond MJ , McCarthy JJ , Sinha M , et al. Aging and microRNA expression in human skeletal muscle: a microarray and bioinformatics analysis . Physiol Genomics . 2011 ; 43 ( 10 ): 595 – 603 . Crossref PubMed Google Scholar
53. Fredriksson K , Tjäder I , Keller P , et al. Dysregulation of mitochondrial dynamics and the muscle transcriptome in ICU patients suffering from sepsis induced multiple organ failure . PLoS One . 2008 ; 3 ( 11 ): e3686 . Crossref PubMed Google Scholar
54. Fearon KC , Voss AC , Hustead DS , Cancer Cachexia Study Group . Definition of cancer cachexia: effect of weight loss, reduced food intake, and systemic inflammation on functional status and prognosis . Am J Clin Nutr . 2006 ; 83 ( 6 ): 1345 – 1350 . Crossref PubMed Google Scholar
55. Chinchilla A , Lozano E , Daimi H , et al. Microrna profiling during mouse ventricular maturation: a role for miR-27 modulating MEF2C expression . Cardiovasc Res . 2011 ; 89 ( 1 ): 98 – 108 . Crossref PubMed Google Scholar
56. Jiang G , Huang C , Li J , et al. Role of STAT3 and FOXO1 in the divergent therapeutic responses of non-metastatic and metastatic bladder cancer cells to miR-145 . Mol Cancer Ther . 2017 ; 16 ( 5 ): 924 – 935 . Crossref PubMed Google Scholar
57. Zimmers TA , Fishel ML , Bonetto A . Stat3 in the systemic inflammation of cancer cachexia . Semin Cell Dev Biol . 2016 ; 54 : 28 – 41 . Crossref PubMed Google Scholar
58. He WA , Calore F , Londhe P , Canella A , Guttridge DC , Croce CM . Microvesicles containing miRNAs promote muscle cell death in cancer cachexia via TLR7 . Proc Natl Acad Sci U S A . 2014 ; 111 ( 12 ): 4525 – 4529 . Crossref PubMed Google Scholar
59. Fearon K , Strasser F , Anker SD , et al. Definition and classification of cancer cachexia: an international consensus . Lancet Oncol . 2011 ; 12 ( 5 ): 489 – 495 . Crossref PubMed Google Scholar
60. Rosenberg IH . Sarcopenia: origins and clinical relevance . J Nutr . 1997 ; 127 ( 5 Suppl ): 990S – 991 . Crossref PubMed Google Scholar
61. Zacharewicz E , Della Gatta P , Reynolds J , et al. Identification of microRNAs linked to regulators of muscle protein synthesis and regeneration in young and old skeletal muscle . PLoS One . 2014 ; 9 ( 12 ): e114009 . Crossref PubMed Google Scholar
62. Zheng Y , Kong J , Li Q , Wang Y , Li J . Role of miRNAs in skeletal muscle aging . Clin Interv Aging . 2018 ; 13 : 2407 – 2419 . Crossref PubMed Google Scholar
63. Reginster J-Y , Beaudart C , Buckinx F , Bruyère O , Osteoporosis BO . Osteoporosis and sarcopenia: two diseases or one? Curr Opin Clin Nutr Metab Care . 2016 ; 19 ( 1 ): 31 – 36 . Google Scholar
64. Materozzi M , Merlotti D , Gennari L , Bianciardi S . The potential role of miRNAs as new biomarkers for osteoporosis . Int J Endocrinol . 2018 ; 2018 : 1 – 10 . Crossref PubMed Google Scholar
65. Goldring MB , Otero M . Inflammation in osteoarthritis . Curr Opin Rheumatol . 2011 ; 23 ( 5 ): 471 – 478 . Crossref PubMed Google Scholar
66. Chen H , Chen L . An integrated analysis of the competing endogenous RNA network and co-expression network revealed seven hub long non-coding RNAs in osteoarthritis . Bone Joint Res . 2020 ; 9 ( 3 ): 90 – 98 . Crossref PubMed Google Scholar
67. Balaskas P , Goljanek-Whysall K , Clegg P , et al. Microrna profiling in cartilage ageing . Int J Genomics . 2017 ; 2017 : 1 – 11 . Crossref PubMed Google Scholar
68. Shorter E , Sannicandro AJ , Poulet B , Goljanek-Whysall K . Skeletal muscle wasting and its relationship with osteoarthritis: a mini-review of mechanisms and current interventions . Curr Rheumatol Rep . 2019 ; 21 ( 8 ): 1 – 8 . Crossref PubMed Google Scholar
69. Soriano-Arroquia A , House L , Tregilgas L , Canty-Laird E , Goljanek-Whysall K . The functional consequences of age-related changes in microRNA expression in skeletal muscle . Biogerontology . 2016 ; 17 ( 3 ): 641 – 654 . Crossref PubMed Google Scholar
70. Nakamura A , Rampersaud YR , Nakamura S , et al. microRNA-181a-5p antisense oligonucleotides attenuate osteoarthritis in facet and knee joints . Ann Rheum Dis . 2019 ; 78 ( 1 ): 111 – 121 . Crossref PubMed Google Scholar
71. Zhang B , Li Y , Yu Y , et al. Microrna-378 promotes Osteogenesis-Angiogenesis coupling in BMMSCs for potential bone regeneration . Anal Cell Pathol . 2018 ; 2018 : 1 – 9 . Crossref PubMed Google Scholar
72. Soares RJ , Cagnin S , Chemello F , et al. Involvement of microRNAs in the regulation of muscle wasting during catabolic conditions . J Biol Chem . 2014 ; 289 ( 32 ): 21909 – 21925 . Crossref PubMed Google Scholar
73. Sondag GR , Haqqi TM . The role of microRNAs and their targets in osteoarthritis . Curr Rheumatol Rep . 2016 ; 18 ( 8 ): 1 – 23 . Crossref PubMed Google Scholar
74. Hanna J , Hossain GS , Kocerha J . The potential for microRNA therapeutics and clinical research . Front Genet . 2019 ; 10 : 1 – 6 . Crossref PubMed Google Scholar
Author contributions
J. Brzeszczyńska: Conceptualized and designed the paper, Reviewed the literature, Drafted and revised the manuscript.
F. Brzeszczyński: Reviewed the literature, Drafted and revised the manuscript.
D. F. Hamilton: Conceptualized and designed the paper, Revised the manuscript.
R. McGregor: Conceptualized and designed the paper, Revised the manuscript.
A. H. R. W. Simpson: Conceptualized and designed the paper, Revised the manuscript.
Funding statement
No benefits in any form have been received or will be received from a commercial party related directly or indirectly to the subject of this article.
ICMJE COI statement
J. Brzeszczyńska reports a fellowship from the Daphne Jackson Trust during the writing of this paper. A. H. R. W. Simpson reports institutional grants from RCUK, Stryker, and is an employee of Bone and Joint Research.
© 2020 Author(s) et al. This is an open-access article distributed under the terms of the Creative Commons Attribution Non-Commercial No Derivatives (CC BY-NC-ND 4.0) licence, which permits the copying and redistribution of the work only, and provided the original author and source are credited. See https://creativecommons.org/licenses/by-nc-nd/4.0/.
